

Epiphytische Flechten

Flechten sind eine Lebensgemeinschaft zwischen Alge(n) und Pilz(en). Wie an-dere Organismen, zeichnen sich Flech-ten durch eine ihnen eigene Farben- und Formenvielfalt aus.
Es gibt eine Fülle an Übersichts-Infor-mationen zu dieser Organismengruppe. So beschränkt sich diese Seite auf spe-zielle Fragen (mehr Fragen als Antwor-ten) zur Ökologie der auf anderen Pflan-zen aufsitzenden - epiphytischen - Flechtenarten.
Kost und Logis

Den Begriff symbiontische Lebensge-meinschaft aus Alge(n) und Pilz(en) verdeutlichen die Querschnitte der Furchen-Schüsselflechte (Blattflechte).
Von oben nach unten:
a) Den oberen Abschluss bildet die obe-
re weiß-graue Rindenschicht.
b) An sie schließt eine grüne, von Pilz-
fäden umsponnene Algenschicht an.
c) Danach folgt eine weiße Markschicht
aus locker wachsende Pilzfäden
d) Es folgt die untere Rindenschicht.
e) Die wie Wurzeln aussehenden dunk-
len Rhizinen haften am Substrat
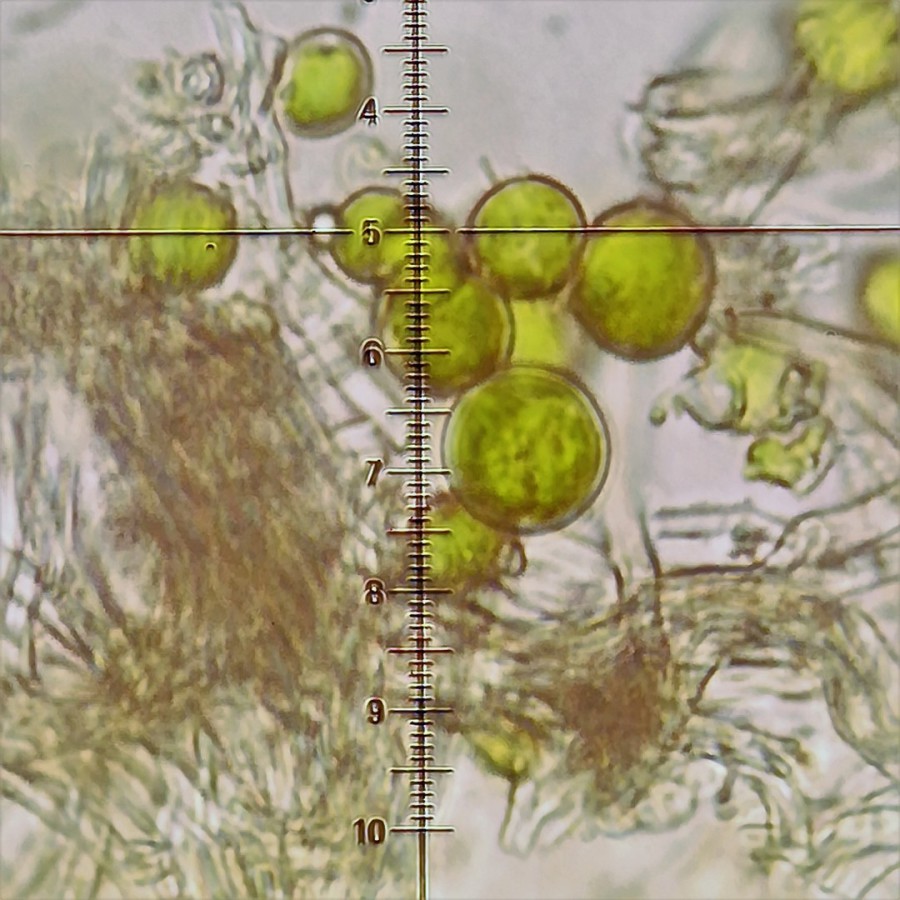
Die in Pilzfäden eingesponnenen Algen
befinden sich in einer Schicht des Flech-tenkörpers oder -lagers. Sie betreiben
Photosynthese, assimilieren und ver-
sorgen den Pilzpartner mit Zuckern und Zuckeralkoholen (Kost), während die Pilzfäden das Lager (Logis) bereitstellen.
In Gallert- und manchen Krustenflechten sind Alge und Pilz eher gleichförmig ver-teilt.

Darüber, warum sie nun Flechten (Liche-nes) heisen, geben etymologische Wör-terbücher wenig her. Etwa soviel man weiß, warum das Haus zur Flechte in der 'Neuen Frankfurter Altstadt' eben diesen Namen trägt.
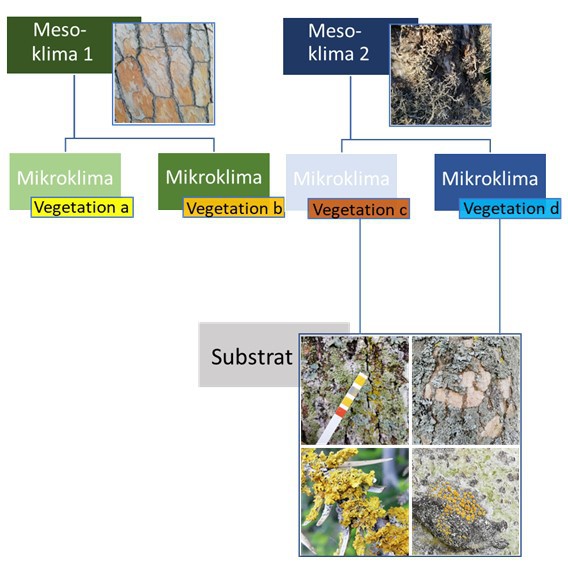
Ökologie
Die Lebensansprüche epiphytischer Flechten und die Komplexität ihrer Umwelt zu entschlüsseln, wird man bislang nur ansatzweise gerecht. Zweifellos wurden zwi-schenzeitlich viele Standortparameter - teils besser (Borken-pH), teils schlechter (Lichtklima im Jahresverlauf) - gemessen und mit dem Flechtenbewuchs (Arten-zusammensetzung, Deckung...) korreliert. An Daten, vor allem solche, die über Einfachmessungen gewonnen werden können, besteht kaum Mangel1.
Das Problem besteht darin, dass zu den Ansprüchen der Flechten(gemeinschaften), ihrer Physiologie und ihren Populationen kaum raum-zeitlich skalierte Modelle, mit einer über die Einzelstudie hinausgehenden Gültigkeit, vorliegen. Die über 60 Jahre alte hierarchische Gliederung wirksamer Umweltfaktoren auf epiphytische Kryptoga-mengemeinschaften (Barkman 1958) ist daher noch heute umfassend und modern.
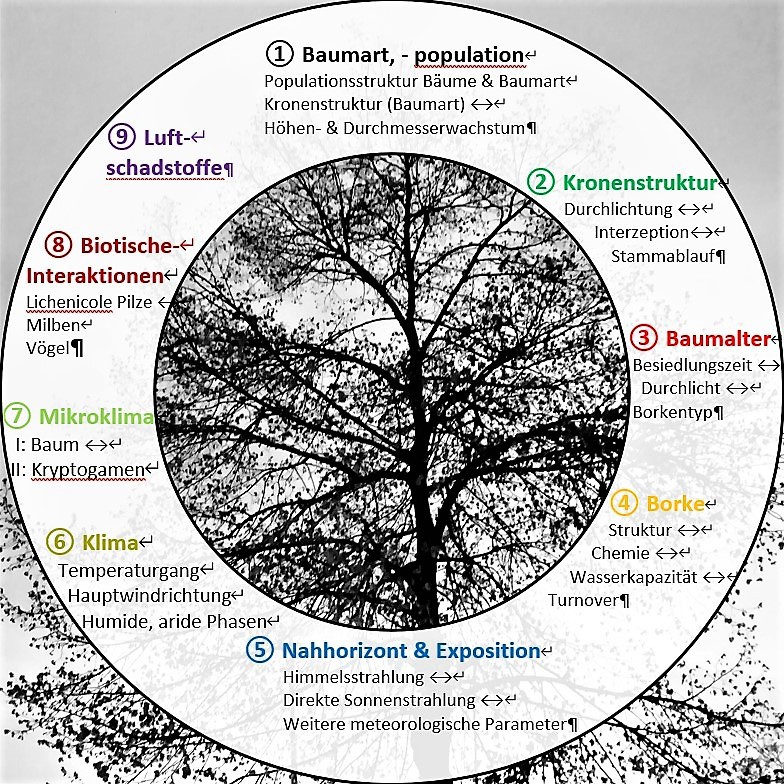 Bioindikation - ein komplexes Unterfangen. Beabsichtigt man etwa Luftschadstoffe/Klimawandel mit Hilfe epi-phytischer Kryptogamen nachzuweisen, wären jeweils 8 weitere Faktoren möglichst konstant zu halten.
Bioindikation - ein komplexes Unterfangen. Beabsichtigt man etwa Luftschadstoffe/Klimawandel mit Hilfe epi-phytischer Kryptogamen nachzuweisen, wären jeweils 8 weitere Faktoren möglichst konstant zu halten.
Inwieweit jeder der Einflussfaktoren
1 - 9 überhaupt relevant, und falls ja, unabhängig (Orthogonalität in Raum und Zeit) von den anderen betrachtet werden kann, wird in den nachfolgen-den Rubriken, wenn nicht geklärt, so doch (anschaulich) hinterfragt.
1. Flechtenzusammensetzung in unterschiedlichen Höhen am Stamm (Räumliche Differenzierung)
An einer mindestens 80jährigen entwurzelten Birke von rd. 25 Meter Höhe, wurden auf jeweils 1 Meter langen Abschnitten (Höhen: 2m, 10m, 20m) die epiphytischen Flechten bestimmt. Da der Ausbreitung der Arten (Soredien, Isidien, Thallusbruch-stücke) von oben nach unten nichts im Wege steht, könnte man vermuten, dass die Arten in 20 Metern Höhe auch im unteren Stammbereich wachsen.





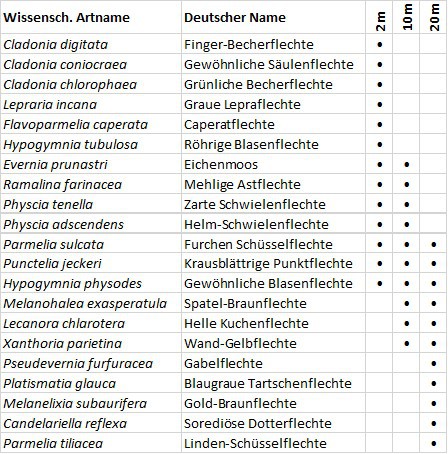






Offenbar tun sie das nicht. Ein Grund dafür könnte sein, dass im Kronenbereich
(s. Bilder) noch die junge weisse Glattrinde (Ringelkork; Funktion von Initialperi-derm ca. 20 bis 40 Jahre) und im unteren Bereich die deutlich ältere netzförmige Schuppenborke vorherrscht.
Diese Borkentypen sind
- unterschiedlich alt (Besiedlungszeitraum. Moose: Konkurrenz & ökologische Förderung2)
- unterschiedlich dynamisch (Borkenturnover erfordert Anpassung)
- unterschiedlich rauh, feuchtehaltend, sauer etc. (Standortunterschiede i.e.S.)
Unabhängig von den Borkeneigenschaften ist das Lichtklima in der Vegetationszeit im Kronenbereich ein anderes als an der laubfreien Basis.1 Die bodennahe 2m-Luft-schicht der Waldlichtung ist hinsichtlich des Wasser-Sättigungsdefizits (Bodennebel) anders zu beurteilen, als in 20 Meter Höhe. Anders ausgedrückt: Nutzen die oberen Flechten(stockwerke) eher Regen, die unteren Nebel/Luftfeuchte? Daneben bestehen noch weitere mikroklimatische Unterschiede zwischen den Wuchsorten.
Die Flechtenvegetation (Einzelfall Birke!) differenzierbarer Baumstockwerke unter-scheidet sich floristisch als auch quantitativ (z.B. auch John & Schroeck 2001). Voraussetzung ist aber zunächst, dass Kandidaten zur Besiedlung der unterschied- lichsten Kleinstlebensräume eines Baumes überhaupt vorhanden sind, was nur für Gegenden mit schwacher Luftbelastung zutrifft. Daneben entsteht Lebensraumviel-falt für Epiphyten nur, wenn Bäume alt werden dürfen.
Die vertikalen Besiedlungsmuster an Birken in South Devon (GB) (Harris 1971) ähneln hinsichtlich Zonierung und Artenzusammensetzung der hier gezeigten Einzel-aufnahme. An insgesamt 25 Birken wurden dabei 25 Flechtenarten nachgewiesen.
Die Reihung der Baumarten nach ihrer Borkentextur (Klassen 1 - 10) und ihrer Güte als Flechtensubstrat (Müller 1981) die junge Birken als eher ungeeignet (2), alte als eher geeignet (9) ausweist, ist allerdings zu hinterfragen, da auch ein alter Stamm im Gipfelbereich Eigenschaften (Ringelkork) und Alter junger Birken aufweist.
Einfluss-Komplex 1: (Baum)art spezifische Variabilität
Einfluss-Komplex 4: Borken-Turnover und der unterschiedliche Besied-
lungszeitraum könnte die Artenzusammensetzung
epiphytischer Flechten beeinflussen
Einfluss-Komplex 3: Die Entwicklung des Einzelbaumes (Ontogenese).
Epilog
(oder: zur Bedeutung einzelner Bäume)



2. Entwicklung epiphytischer Flechten im unteren Stammbereich (Zeitliche Differenzierung)
Die Bilder zeigen drei in 50 Meter Umkreis wachsende Sommerlinden mit Stamm-
durchmessern von 8, 28 und 150 Zentimetern. Eine mögliche Sukzessionsreihe ausgehend von Krustenflechten über kleinlappige Blattflechten, bis hin zu einer Vielfalt der Wuchsformen.








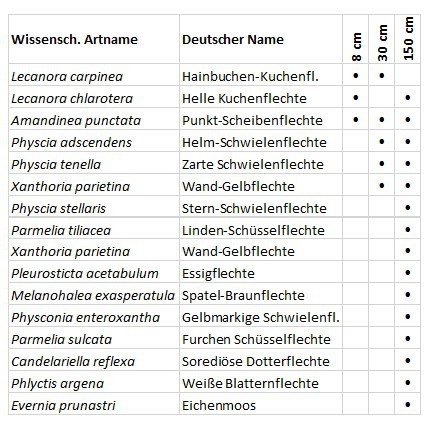
Einfluss-Komplex 3: Das Baumalter steht in Zusammenhang mit
a) der Wahrscheinlichkeit den Stamm zu besiedeln (Zeit)
b) der Änderung der Rindenstruktur (und Kronenstruktur)
Glatte Rinde → streifenförmige Borkenrisse → Netzborke



c) (chemischen Eigenschaften)
3. Licht- und Feuchteverhältnisse am Stamm
Hale (1983) postuliert als wichtige Faktorenkomplexe, die die Artengemeinschaft epiphytischer Flechten differenzieren, die Substratfaktoren (60%) und das Mikro-klima (40%). Nach Wirth & al. (2013) ist die „Baumartenbindung der Flechten mit wenigen Ausnahmen gering.“ Nur eine Einteilung in basenreiche und basenarme Rindenoberflächen scheint flechtenökologisch bedeutsam zu sein.
Ohne weiteres ließen sich solche Expertenaussagen als Nullmodell oder Grundan-nahmen für mittlere Breiten bei fehlendem Immissionseinfluss beibehalten. Schnell kommt man bei der Beschreibung regionaler und lokaler Sachverhalte damit an Grenzen. Schon allein deshalb, weil das Mikroklima und daher auch eine mikroklima-tisch bedeutsam erachtete Struktur - z.B. Borken(riss)tiefe - durch das Mesoklima beeinflusst wird und daher einen Bedeutungswandel (Relative Standortkonstanz) erfährt. Anders ausgedrückt, selbst (absolut) gemessene Standortfaktoren haben auf Organismen eine nur relative Bedeutung.
Da Lebensgemeinschaften nicht ausschließlich durch abiotische Randbedingungen geprägt werden, sondern maßgeblich auch durch populationsbiologisch wirksame Prozesse, wie dem Diasporenaustausch, ist auch dieser in irgendeiner Form im Aufnahmeumfeld der Flechten zu berücksichtigen.






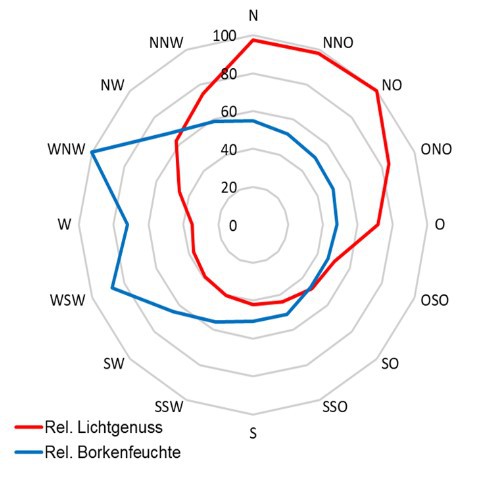
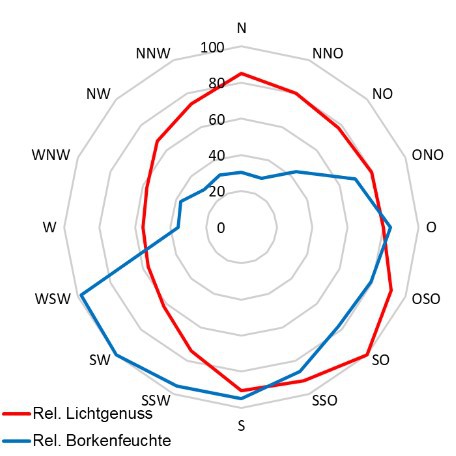



Beispiel wäre der Epiphytenbewuchs einer best. Baumart auf Meereshöhe und im Hochschwarzwald, oder die Lebensverhältnisse der Flechten in einer Stadt in Deutschland mit 20, oder einer anderen mit 80 Nebeltagen.
Limnoterra ist - auch international -
keine Studie bekannt in der Feuchte-
anteile (Niederschlag, Nebel, Luft-feuchte) in ihrer Bedeutung bestimmt und in Relation zueinander und auf die epiphytische Flechtenvegetation gesetzt wurden.
Allgemein ausgedrückt:
Der Stellenwert lokal oder regional als ökologisch bedeutend erachteter Standort-parameter kann sich in Abhängigkeit übergeordneter klimatischer Randbedingungen ändern.
Ein hierarchisches System3, auf Grund dessen entschieden werden kann, welche Ein-flussfaktoren bei gegebener Geländesituation als konditional, operational und als re-sultierend anzusehen sind, wäre zur Einordnung zahlreicher ökologischer Studien
- die nach Betrachtungsweise und Messprogramm einen anderen Faktor zum Schlüs-selfaktor erhebt3 - definitiv zielführend.
Einfluß-Komplex 5
Einfluß-Komplex 2: Insbesondere im besiedelten Bereich spielen einengende Struk-
turen wie Gebäude und Mauern, aber auch andere Bäume eine große Bedeutung für die mögliche tägliche Besonnungsdauer.
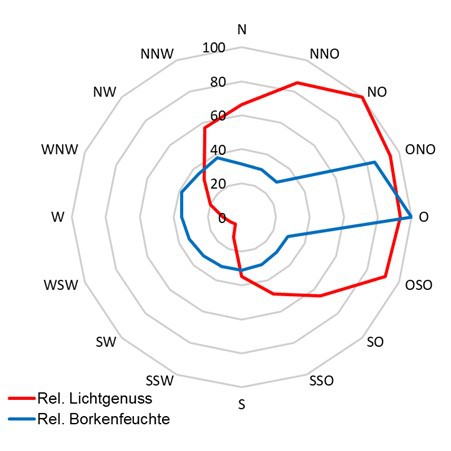
Die Kronenstruktur spielt nicht nur hinsichtlich der Belichtungs-Situation eine Rolle, sondern sie beeinflusst darüber hinaus Stammablauf, Kronendurchlässigkeit und damit die Benetzung der Flechten.
Dies geht einher mit der Substratfeuchte, auch wenn diese nicht zu den primär wichtigen Standortfaktoren (Wirth 1992) gehören mag.
Da Licht (Wärme) bei Flechten nur zusammen mit Feuchte
- nur bei feuchtem Thallus sind Flechten photosynthetisch aktiv - ökologisch relevant ist, ist es günstig, beides einer gemeinsamen Betrachtung zu unterziehen. Die Befeuchtung der Flechten kann
über Regen, Tau und Nebel erfolgen, was die standörtliche Beur-teilung weiter kompliziert.
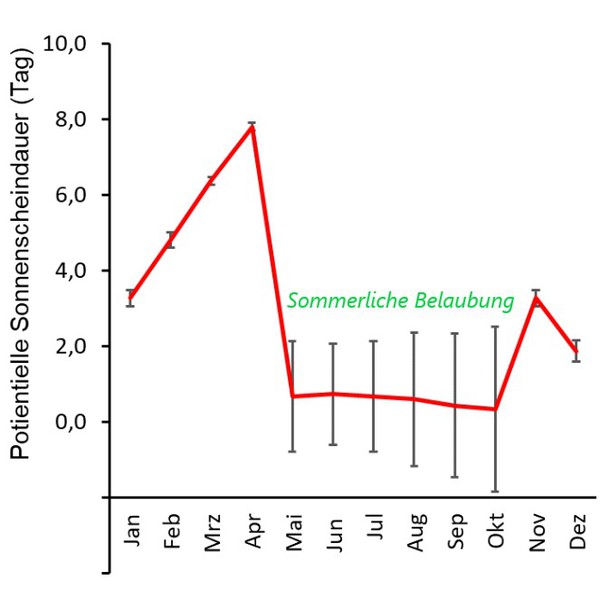
Flechten sind lichtliebende Organismen, dennoch wird selbst bei wissenschaftlichen Studien das Lichtklima oft gar nicht, oder unzureichend
erfasst. Im Gegensatz zu einer einmaligen pH-Messung müssen zur Beurteilung des Lichtklimas am Stamm wenigstens zwei Messungen - im belaubten und unbelaubten Zustand - erfolgen. In jedem Fall ist die
Kenntnis der Potentiellen Sonnenscheindauer, oder wenigstens des Sky View Factors zielführend, um lichtklimatisch bedeutsame Strukturen (Nahhori-zont) in die Beurteilung einzubeziehen.
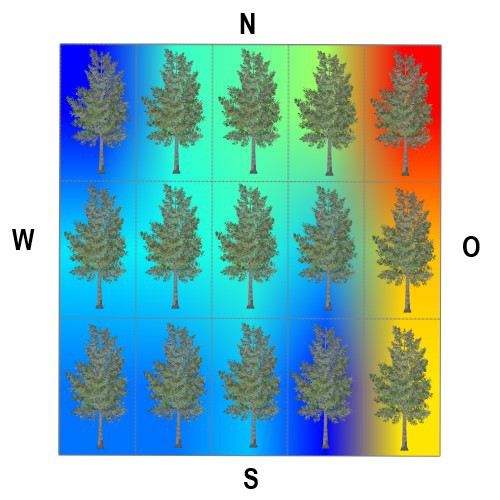
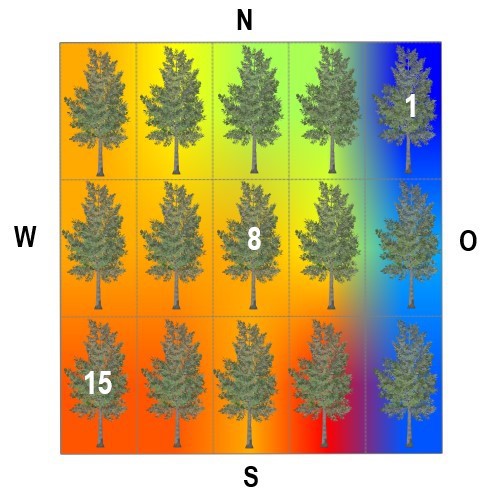
Die Bedeutung der Stammexposition ist, angesichts der Wirkung von diffuser Him-melsstrahlung, sommerlicher Belaubung, der Wechselwirkung Licht/Feuchte und
ggf. der 'Streßwirkung Südexposition', nicht eindeutig.
Astoberseiten stellen praktisch immer günstigere Wuchsbedingungen für viele epi-
phytische Generalisten unter den Flechten bereit.
4. Epiphytische Flechten als Nahrung
Es gibt Pilze, die Algen parasitieren - sowohl freilebende als auch die symbiontisch lebenden Grün- und Blaualgen der Flechten. Einer dieser Flechtenparasiten ist der in Deutschland weit
verbreitete Weiße Rindenpilz, auch Große Algenspinne/Zweisporige
Gewebehaut (Athelia arachnoidea) genannt.
Wird es auf einem Baum allzu bunt, sind u.U. mehrere solcher Gegenspieler der Flechten mit von der Partie.







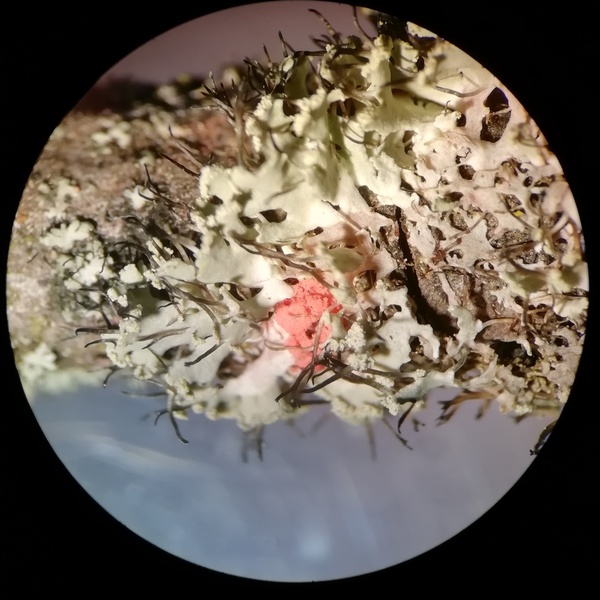







Einfluss-Komplex 8: Lichenicole Pilze haben zumindest lokal einen differenzierenden
Einfluss auf die epiphytische Flechtenvegetation, was bei immis-
sionsökologischen Untersuchungen zu berücksichtigen ist.
5. Zur Substratspezifität epiphytischer Flechten
Im Gelände
In den feuchten Tropen können bestimmte Flechten fast jedes Substrat in kürzester Zeit besiedeln. Im temperaten Klima Deutschlands bedarf es meist längerer Zeit-räume, bis ubiquitäre Arten schwer besiedelbare Substrate einnehmen. Vor allem spielen dabei die Ausbreitungsverhältnisse eine Rolle (z.B. werden Oberflächen von Kabelverteilerschränken unter flechtenreichen Bäumen schnell bewachsen).
„Floristisch heben sich diese selten besiedeten Materialien jedoch nicht hervor“, wie Wirth (1976) konstatiert. Hinsichtlich der Spezifität hat die Aussage immer noch Gültigkeit, während von ausgesprochener Seltenheit kaum mehr die Rede sein kann.
Da wir Pflanzen-verwertbaren Stickstoff in ausreichender Menge in die Luft blasen, erfolgt die Ansiedlung u.U. innerhalb weniger Jahre. Auch Kunststoff- und Glasab-deckungen von Photovoltaik-Anlagen werden von Flechten (Algen, Moosen) besie-delt, was u.a. zur Entmineralisierung der Gläser führt.
Flechten-Entfernung: ein lukratives Geschäftsmodell der nächsten Jahrzehnte?










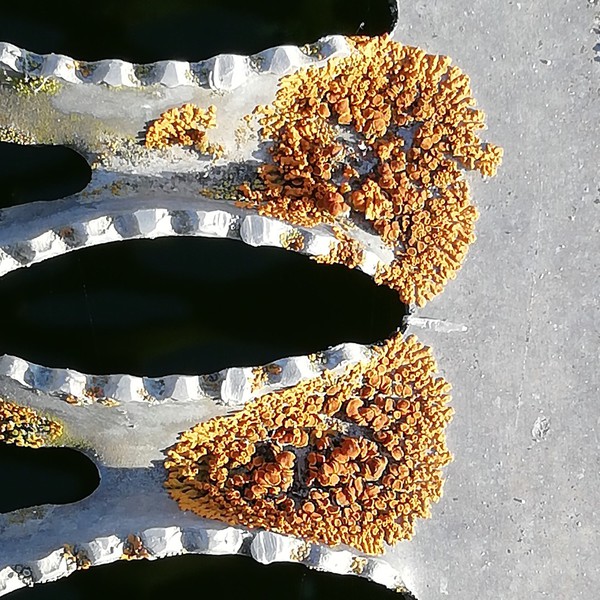



Einfluss-Komplex 4: Wenigstens die
Kunststoff-Substrate stellen epiphytischen
Flechten praktisch keine chemischen Ressourcen bereit.
Hinsichtlich des oberflächigen Säurewertes sind sie neutral.
Auf solchen Substraten siedelnde Flechten leben ausschließ-
lich von Beaufschlagungen
der Atmosphäre (Wasser, Mineral-
stoffe), CO2 und Sonnenlicht.
Kunstsubstrat-Exposition epiphytischer Flechten
Mikroklima, Substratfaktoren und Luftnähr-/schadstoffe – diese Faktoren bestimmen das Wachstum aller Flechtenarten. Der am natürlichen Wuchsort jeweilig herrschen-
de Wirkungsanteil ist zwar schon mehrmals beziffert worden, muss aber wohl ab-hängig von Flechtenart und spezifischem Standort jeweils neu austariert werden.
Um die durch unterschiedliche Substrate hervorgerufene Variabilität zu egalisieren, bietet sich bei der Exposition von Flechten - etwa zum Nachweis von Luftschadstof-fen - die Verwendung einheitlicher künstlicher Substrate an. Hierbei können unter-schiedliche Flechtenarten mit ausreichender Wiederholungszahl (Auswerte-Statistik) exponiert werden.






Die Flechten bzw. einzelne Flechtenloben werden auf Deckeln von Petrischalen mit 55 mm Durchmesser exponiert. Zur Befestigung werden sie auf eine lackierte Reiß-zwecke (hier sind bessere Lösungen aus Plastik wünschenswert!) aufgesteckt.
Diese wurde zuvor mit einem doppelseitigen Klebeband mittig auf den Deckel aufge-klebt und darauf ein passgenauer Glasfaserrundfilter aus Borosilikat-Glasmikrofaser (ohne Bindemittel) aufgebracht.
Die Petrischalen werden in die Auflagedeckel von Eurobehältern eingepasst. Durch deren Profilrand lassen sich die Flechten mit straffer Nylongaze als Schutz vor Vö-geln (Flechten als Nistmaterial) sowie Regen- und Hagelschlag schützen.
Die Wachstumsanalyse erfolgt mit Hilfe von Bildanalyse-Software wie ImageJ.4
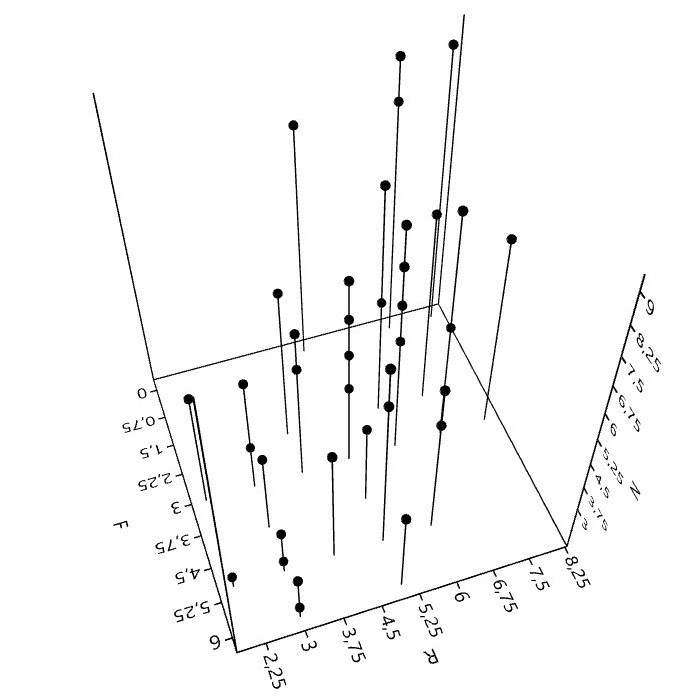
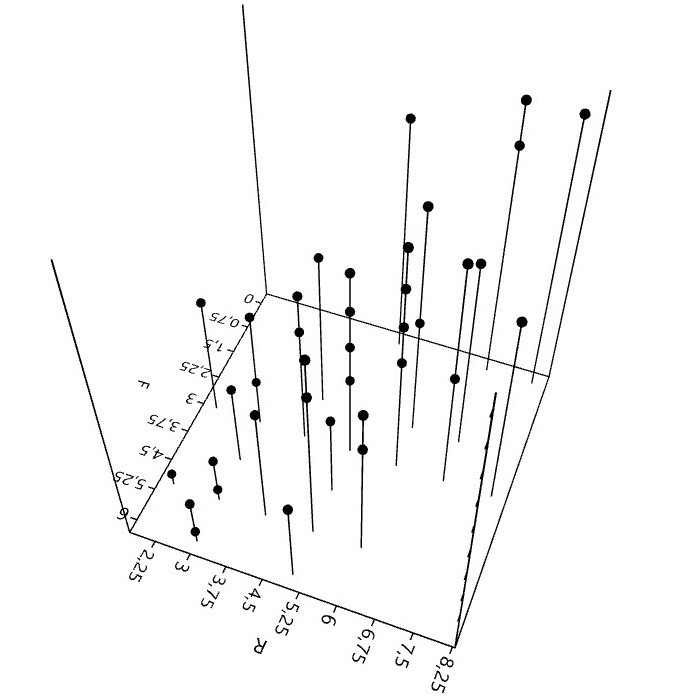
6. Mit Flechten rechnen (Zeigerwerte)
Bei der Kartierung epiphytischer Flechten in Städten, spielen die 50 aufgeführten Arten eine wichtige Rolle. Sie bilden hinsichtlich Präsenz und Häufigkeit derzeit etwa das Basisartengefüge. 70 bis 80% der gesamten Artenzahl, Frequenzsumme, Deckung oder ein anderes Bedeutungsmaß entfällt auf diese Arten.
Bei Verwendung der (Arten)Zeigerwerte5 und der Interpretation daraus berechneter Bestandeszeigerwerte, ist kritische Distanz nötig.
 in Städten.")
.")
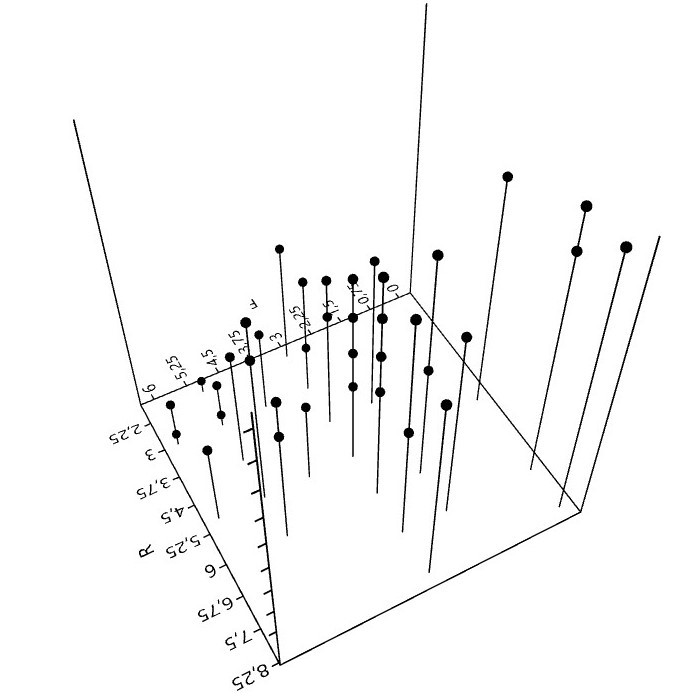
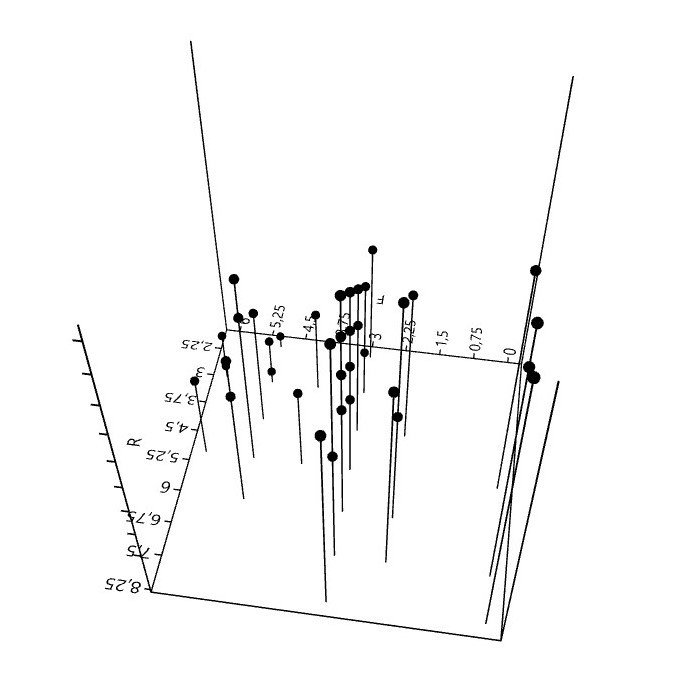
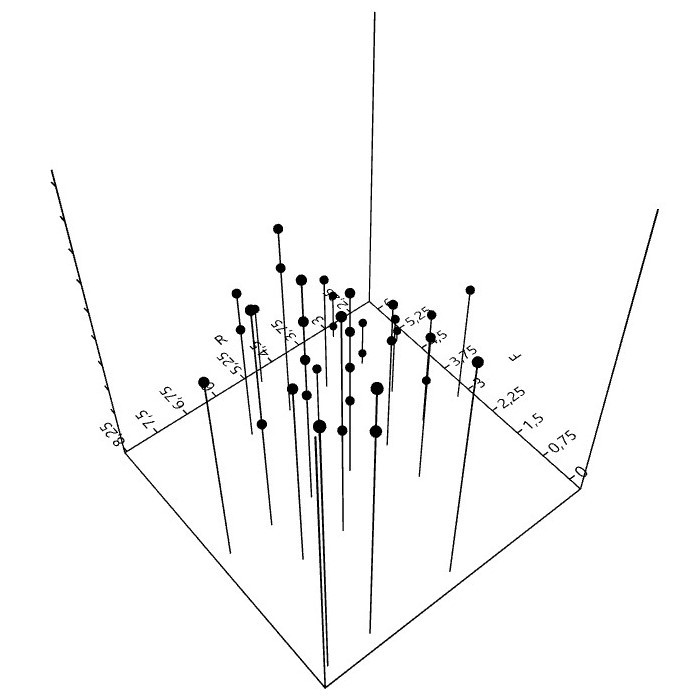
 der Zeigerwerte.")
Warum?
Zeigerwerte sind nicht unabhängig. Für sechs, auf Grundlage der genannten 50 Basisarten berechneten Zeigerwerte, lassen sich paarweise 15 Rangkorrelationen berechnen. Für Feuchte, Reaktion und die Eutrophierungszahl (N) sind die Korre-lationen hochsignifikant.6
Daher stellt sich die Frage, ist mit der Angabe eines Bestandeszeigerwerts (F, R, N) der jeweilige Umweltfaktor wirklich wissenschaftlich zu erschließen, oder müsste hierzu eine stärkere Unabhängigkeit der Zeigerwerte, was vielleicht nicht in ihrer 'Natur' liegt, bestehen?
Auch wenn bestimmte Arten heute auf Grund von Experteneinschätzungen (Zeiger-werte) als Indikatoren für Klimawandel (eine jegliche Vegetation indiziert auch Kli-mawandel) präferiert werden, ist damit (s. Kapitel 3) nicht geklärt, welches nun die meteorologisch wirksamen Anteile (bzw. deren Wechselwirkungen) sind, die der be-obachteten Arealverschiebung zugrunde liegt.
Die Erhöhung der Jahresmittel-Temperatur kann angesichts der Temperaturtoleranz der Flechten im trockenen Zustand, kaum Schlüssel-Parameter sein. So lassen sich zwar Veränderungen beobachten, kaum allerdings eine Erweiterung unserer Kennt-nisse.
1
Flechten sind Lichtpflanzen. Wachstum ist aber nur bei ausreichender Thallusfeuchte
möglich. Der Zu-stand 'ausreichend Licht (Wärme) + ausreichend Feuchte' ist am Stamm-Lebensraum von geringerer Eintrittswahrscheinlichkeit als beispielweise im
Kronenraum. Auch müsste die EW wenigstens für die Jahreszeiten differenziert beziffert werden. Singulär betrachtete Mehrjahres-Mittelwerte (Klimahüllen) oder kurzfristig erhobene Faktoren
("Witterungs-Hüllen") skizzieren daher nur grobe Randbedingungen, die bei der Interpretation des Einzelfalls u.U. mehr verschleiern, als erhellen.
Die Tatsache, dass der Kronenbereich den in der Regel kartierten unteren Stammbereich mit Ausbrei-tungseinheiten 'füttert' (siehe aber Bsp. 1), macht das Ganze nicht einfacher.
2
Hier: Dicranum scoparium, Dicranoweisia cirrata, Hypnum cupressiforme, Orthotrichum spec..
Gefördert werden beispielsweise Flechten der Gattung Cladonia auf
Moos-Substrat.
3
Barkman, J. J. (1958): On the Ecology of cryptogamic epiphytes. With special reference
to the Netherlands. Proefschrift. Rijksuniversiteit te Leiden. 202 S.. Z.B. Unterkapitel 16.
Für allen Freiland-ökologischen Studien gilt, dass die Parameterwahl plausibel sein mag, deren Messung letztlich immer den zeitlichen, finanziellen und technischen Möglichkeiten geschuldet ist.
Eine Einschätzung der Bedeutung erhobener Standortfaktoren hinsichtlich Vorkommen und Wachstum
der Epiphyten erfolgt dann meist a posteriori durch Korrelationsmaße, die nicht
zwangsläufig eine Kau-salbeziehung wiedergeben. Ob weitere, nicht einbezogene Faktoren in stärkerem Maße Koinzidenz auf-weisen, bleibt im Dunkeln.
So sind Daten zur Jahresmitteltemperatur einfach zu generieren und werden in kaum einem Vergleich von Wuchsorten fehlen. Hingegen ist ein Standortsvergleich nach Häufigkeit und Zeitpunkt der Taubil-dung in höherem Maße bedeutsam für Epiphytenvorkommen, etwa in Städten, doch wird man hierzu kaum wissenschaftliche Untersuchungen finden. Für eine Veröffentlichung reichen meist ubiqitäre Da-ten. Parameter, die eine Hypothesenbildung ideal stützen würden fehlen oft, weil sie aufwändig zu er-heben sind. So entwickeln wir durch unser vorherrschend korrelatives Denken zwar Vorstellungen was sein könnte, wissen aber nicht, was tatsächlich passiert.
4
Tremp, H. (2023): Das Wachstum von Randloben epiphytischer Flechten auf Kunstsubstrat - Methodische Vorarbeiten für ein erweitertes Biomonitoring. Gefahrstoffe 85(5-6). 85 - 90.
5
Ellenberg, H., Weber, H. E., Düll, R., Wirth, V., Werner, W., & Paulißen, D. (1992): Zeigerwerte von Pflanzen in Mitteleuropa. Datenbank. Scripta Geobot, 18, 1 - 258.
Wirth, V. (2010): Ökologische Zeigerwerte von Flechten - erweiterte und aktualisierte Fassung.
Herzogia, 23(2), 229-248.
6
„It seems that Wirth's N scale, at least for epiphytic species, runs more or
less parallel with the
R scale.“
van Dobben, H. F. & C.J.F. ter Braak (1999): Ranking epiphytic lichen sensivity to air pollution using survey data: a comparison of indicator scales. Lichenologist
31(1). 27 - 39.